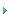要比a1和(A)之间的差异为大。假定m1产生两个变种,即 m2和s2,彼此不同,而和它们的共同亲代(A)之间的差异更大。我们可以用同样的步骤 把这一过程延长到任何久远的期间;有些变种,在每一千代之后,只产生一个变种,但 在变异愈来愈大的条件下,有些会产生两个或三个变种,并且有些不能产生变种。因此 变种,即共同亲代(A)的变异了的后代,一般会继续增加它们的数量,并且继续在性状 上进行分歧,在图表中,这个过程表示到一万代为止,在压缩和简单化的形式下,则到 一万四千代为止。
但我在这里必须说明:我并非假定这种过程会像图表中那样有规则地进行(虽然图 表本身已多少有些不规则性),它的进行不是很规则的,而且也不是连续的,而更可能 的是:每一类型在一个长时期内保持不变,然后才又发生变异。我也没有假定,最分歧 的变种必然会被保存下来:一个中间类型也许能够长期存续,或者可能、也许不可能产 生一个以上的变异了的后代:因为自然选择常常按照未被其他生物占据的或未被完全占 据的地位的性质而发生作用;而这一点又依无限复杂的关系来决定。但是,按照一般的 规律,任何一个物种的后代,在构造上愈分歧,愈能占据更多的地方,并且它们的变异 了的后代也愈能增加。在我们的图表里,系统线在有规则的间隔内中断了,在那里标以 小写数目字,小写数目字标志着连续的类型,这些类型已充分变得不同,足可以被列为 变种。但这样的中断是想像的,可以插入任何地方,只要间隔的长度允许相当分歧变异 量得以积累,就能这样。
因为从一个普通的、分布广的、属于一个大属的物种产生出来的一切变异了的后代, 常常会共同承继那些使亲代在生活中得以成功的优点,所以一般地它们既能增多数量, 也能在性状上进行分歧:这在图表中由(A)分出的数条虚线表示出来了。从(A)产生 的变异了的后代,以及系统线上更高度改进的分枝,往往会占据较早的和改进较少的分 枝的地位,因而把它们毁灭;这在图表中由几条较低的没有达到上面横线的分枝来表明。 在某些情形里,无疑地,变异过程只限于一枝系统线,这样,虽然分歧变异在量上扩大 了,但变异了的后代在数量上并未增加。如果把图表里从(A)出发的各线都去掉,只留 a1到a10的那一支,便可表示出这种情形,英国的赛跑马和英国的向导狗与此相似,它们 的性状显然从原种缓慢地分歧,既没有分出任何新枝,也没有分出任何新族。
经过一万代后,假定(A)种产生了a10、f10和m10三个类型,由于它们经过历代性 状的分歧,相互之间及与共同祖代之间的区别将会很大,但可能并不相等。如果我们假 定图表中两条横线间的变化量极其微小,那未这三个类型也许还只是十分显著的变种; 但我们只要假定这变化过程在步骤上较多或在量上较大,就可以把这三个类型变为可疑 的物种或者至少变为明确的物种。因此,这张图表表明了由区别变种的较小差异,升至 区别物种的较大差异的各个步骤。把同样过程延续更多世代(如压缩了的和简化了的图 表所示),我们便得到了八个物种,系用小写字母a14到m14所表示,所有这些物种都是 从(A)传衍下来的。因而如我所相信的,物种增多了,属便形成了。
在大属里,发生变异的物种大概总在一个以上。在图表里,我假定第二个物种(1) 以相似的步骤,经过一万世代以后,产生了两个显著的变种或是两个物种(w10和z10), 它们究系变种或是物种,要根据横线间所表示的假定变化量来决定。一万四千世代后, 假定六个新物种n14到z14产生了。在任何一个属里,性状已彼此很不相同的物种,一般 会产生出最大数量的变异了的后代;因为它们在自然组成中拥有最好的机会来占有新的 和广泛不同的地方:所以在图表里,我选取极端物种(A)与近极端物种(I),作为变 异最大的和已经产生了新变种和新物种的物种。原属里的其他九个物种(用大写字母表 示的),在长久的但不相等的时期内,可能继续传下不变化的后代;这在图表里是用不 等长的向上虚线来表示的。
但在变异过程中,如图表中所表示的那样,另一原理,即绝灭的原理,也起重要的 作用。因为在每一处充满生物的地方,自然选择的作用必然在于选取那些在生活斗争中 比其他类型更为有利的类型,任何一个物种的改进了的后代经常有一种倾向:在系统的 每一阶段中,把它们的先驱者以及它们的原始祖代驱逐出去和消灭掉。必须记住,在习 性、体质和构造方面彼此最相近的那些类型之间,斗争一般最为剧烈。因此,介于较早 的和较晚的状态之间的中间类型(即介于同种中改进较少的和改良较多的状态之间的中 间类型)以及原始亲种本身,一般都有绝灭的倾向,系统线
<< 上一页 [11] [12] [13] [14] [15] [16] [17] [18] [19] 下一页
《物种起源》下载 www.ruida.org/Soft/book/7294.shtml